
Selektionsschatten
Mit Selektionsschatten wird das Nachlassen der Wirksamkeit der natürlichen Selektion (Selektionsdruck) im Verlauf der Lebensspanne einer Spezies bezeichnet. Dahinter steht die Erkenntnis, dass die Reproduktionsphase zwangsläufig auch ohne Altern allein aufgrund von Unglücksfällen wie Blitzschlag, Gefressenwerden usw. endet – die Selektion kommt zum Erliegen. Nur Gene rechtzeitig fortpflanzender Artgenossen gelangen folglich in den Genpool nächster Generationen.Mithilfe des Selektionsschattens wird nun das Altern erklärt. Wenn schon in jungen Jahren schadhafte Genmutationen erfolgen, die aber erst nach Beendigung der im Endeffekt unfallbedingt begrenzten Fortpflanzungsphase zu einem Krankheitsausbruch führen, werden diese nicht evolutionär ausgesiebt. Klassisches Beispiel ist die erbliche, auf einem Gendefekt beruhende Huntington-Krankheit, die erst um das 40. Lebensjahr zu ersten Symptomen führt. Hieraus leitete im Jahr 1942 erstmalig J. B. S. Haldane den evolutionären Mechanismus hinter dem Altern ab. Der Begriff Selektionsschatten wurde indessen durch P. B. Medawar eingeführt, der auch Haldanes Beispiel 1952 zur Mutations-Akkumulations-Theorie verallgemeinerte.
In das Konzept fügt sich die 1957 durch George C. Williams aufgestellte Hypothese der antagonistischen Pleiotropie (so später durch Michael R. Rose benannt), die annimmt, dass manche Gene die Fertilität in jungen Jahren auf Kosten des späteren Überlebens stärken. So ist ein hoher Sexualhormonspiegel fortpflanzungsdienlich, aber auf lange Sicht krebsförderlich. Genauso stütz die 1977 formulierte Disposable-Soma-Theorie von Tom Kirkwood die Annahme eines Selektionsschattens. Weil die evolvierte Fortpflanzungsphase das Überleben des Somas erfordert, werden dorthin Energie allokierende Gene selektiert. Hiernach ist weiters überleben, da nun schon die Genweitergabe gesichert ist, entbehrlich (disponibel), weshalb die Energieallokationen auf Dauer unzureichend sind, so dass erst spät wirksam werdende Schäden akkumulieren können („error catastrophe“).
Das Nachlassen der Selektion ist ein schleichender Prozess, der bald nach Beginn der Geschlechtsreife einsetzt, wenn manche Individuen unfallbedingt keinen weiteren Paarungserfolg haben, und mit dem letztmöglichen aufhört. W. D. Hamilton formalisierte erstmalig 1966 die Zusammenhänge mathematisch nach folgendem Schema:
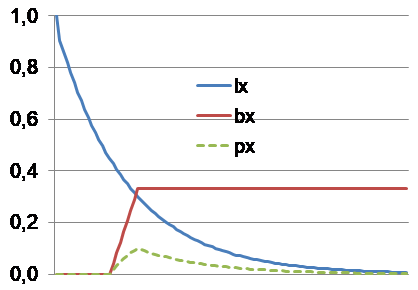
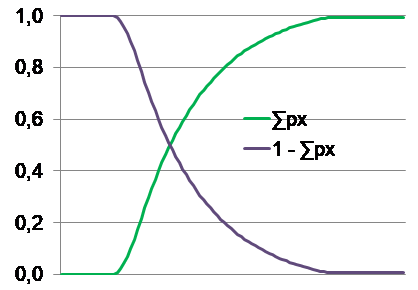
Seien x das chronologische Alter, μ die konstante Dezimierungsrate in der Wildnis (ohne Altern) je vollendetem x, und lx die schwindende Überlebenschance, dann gilt lx = e-μx. Unter Hinzunahme der konstanten Fortpflanzungsstärke bx eines noch überlebenden Individuums lässt sich der erwartbare Fortpflanzungsbeitrag px berechnen: px = lxbx. Die Summe aller Beiträge bis x (sei dies ∑0 bis x genannt) ergibt den wahrscheinlichen gesamten Fortpflanzungserfolg als Maßstab der Fitness. Beim plausiblen „Selbstersatz“, also stabiler Population, wird ∑0 bis ∞ = 1 ergeben. Hierfür muss die Fortpflanzungsstärke hinreichend groß sein. Populationswachstum (∑ > 1) und Schrumpfung (∑ < 1) sind natürlich auch möglich. Für ∑0 bis ∞ = 1 repräsentiert 1 - ∑0 bis x nun den Verlauf der nachlassenden Selektionsstärke, wenn diese anfänglich als 100 % angenommen wird. Die obigen Grafiken mit dem Zeitverlauf auf der x-Achse und der Selektionswirkung auf der y-Achse fassen das Gesagte zusammen.
Verfasser: Dipl.-BW (FH) Michael Zabawa
Erschienen: April 2019
Weiterführende
Literatur


Evolutionäre Medizin: Eine Einführung für Mediziner und Biologen (essentials)

Evolutionäre Medizin: Eine Einführung für Mediziner und Biologen (essentials)